Introduction#
We have all seen or can imagine a frog lashes out its tongue instinctively to capture a flying insect as it approaches. The question that I find fascinating is during the time when the frog (in this case, the predator) was still- was he simply waiting or was he covertly preparing to attack? Although, it is in the nature of the predator to catch the prey, it is good to have some degree of flexibility in behavior, particularly if the prey turns out to be a poisonous variant of the prey species. Similar to this example, in our everyday life we make decisions and prepare ourselves to execute different plans. The hallmark feature of an intelligent behavior is that we can couple or decouple these mental processes from executing them, depending on different circumstances, situations and context. The focus of the thesis is to understand aspects of motor planning and its transformation into a motor action, using eye and hand movements under different task contexts.
Neural Correlates of motor planning#
A wide network of different brain regions are involved in carrying out sensorimotor transformations that are essential towards making a goal directed eye or hand movements.
Early visual areas in the striatal cortex, V1 and V2 are thought to encode different properties like orientation, colour, form and spatial frequency of an object (Campbell et al. 1969, Hubel & Wiesel 1959, Livingstone & Hubel 1988, Maffei & Fiorentini 1973, Sincich & Horton 2005). This information is passed on through the dorsal pathway to different regions in the parietal and frontal areas of the brain that transform visual signals into motor plans. Consistent with this view, the lateral intra parietal (LIP) and parietal reach region (PRR), show effector specific signals (Andersen & Buneo 2002) (but see Bisley & Goldberg 2010, Colby & Goldberg 1999 for a different view). They encode for the spatial location of the target, motor plans for saccadic eye and reach movements, respectively. Also, LIP has been shown to play an important role in saccade decision-making. The signals in these areas are thought to be modulated by high level cognitive functions such as attention which included bottom-up features like luminance, contrast, motion strength (Assad & Maunsell 1995, Balan & Gottlieb 2006, Bisley & Goldberg 2006, Kusunoki et al. 2000) and top-down factors such as reward, value, goal, biases and other intentional states (Ipata et al. 2006a,b; Kusunoki et al. 2000, Platt & Glimcher 1999, Sugrue et al. 2004). LIP projects to frontal eye fields (FEF) and supplementary eye fields (SEF). FEF is thought to play an important role in saccade target selection, response preparation and control of goal directed eye movements (Hanes & Schall 1996, Schall et al. 1995, Schall & Thompson 1999a, Schall 2001). SEF on the other hand has a more executive role in saccade initiation by indirectly controlling the activity of FEF based on prior experience (Stuphorn et al. 2010). The FEF has direct and indirect connections via basal ganglia (BG) to a sub-cortical structure called superior colliculus (SC) in the mid brain. SC has a topographic map which represents saccade vectors (amplitude and direction) that could direct gaze to specified location in space. BG plays an important role in selecting appropriate motor plans and inhibiting irrelevant ones by exerting strong tonic inhibition (Hikosaka & Wurtz 1983) through substantia nigra pars reticulata (SNr), which is one of the output nuclei of BG on SC. SC then projects to saccade generators in pons and midbrain regions which decomposes the saccade vector into horizontal and vertical components before effecting appropriate oculomotor muscle groups to initiate eye movements. In contrast, hand movements are planned in the parietal reach region (PRR) and the anterior intra-parietal region (AIP) that feeds information to the dorsal and ventral premotor cortex (PMd; PMv), respectively. Both these areas are thought to play an important role to reach and grasp objects, respectively. The dorsal premotor area (PMd) is also involved in planning hand movements and thought to encode different kinematic parameters such as direction, amplitude and velocity of an upcoming movement (Churchland et al. 2006), in addition to reach target selection (Song & McPeek 2008). The PMd and PMv, project to motor cortex (M1) for generating motor commands that move the limbs, which in turn drives the motor neurons in the spinal circuitry to cause movements by generating the force required through contraction of appropriate muscle groups. Recent studies also suggest that local processing of information might be going on at each segmental spinal level which not only receive major excitatory inputs from M1 but also has indirect inhibitory projections from premotor areas via the reticulate formation in midbrain (Cohen et al. 2010). Different cognitive processes like decision-making and response preparation have been extensively studied in the context of eye and hand movements. Behavioural studies have suggested that a fair proportion of response time (time to react to a stimuli), could be attributed to the processing of these different central processes. Computationally, these central processes that constitute motor planning can be well described by theoretical frameworks such as accumulation to a fixed bound (Gold & Shadlen 2007). The basis of such accumulation which reflects neural basis for decision making and motor planning are described below. Both these findings are based on studies done in macaques involved in making saccadic eye movements.
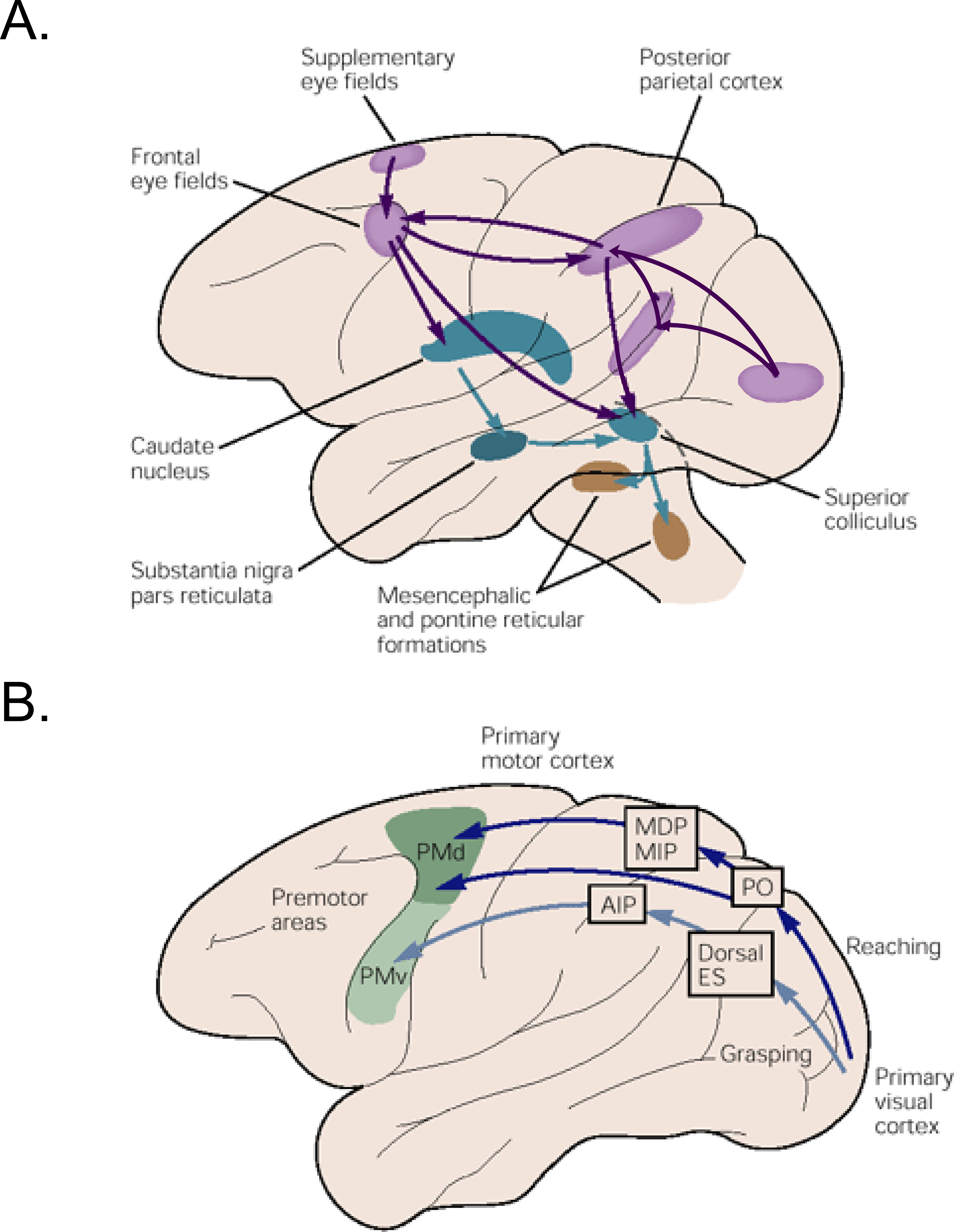
Figure 1.1 Pathways for saccadic eye and hand movement in the macaque brain.#
A) The cortical (pink), sub-cortical (blue) and mid brain regions (brown) are thought to be involved in goal directed saccadic eye movements. Frontal eye field is one of the important oculomotor structures that plays an important role in initiating saccades by directly exciting the colliculus and indirectly releasing it from suppression imposed by the substantia nigra in the basal ganglia. B) Separate cortical regions in the parietal and premotor areas are involved in carrying out visuomotor transformations for reaching and grasping objects. Figures taken from (Kandel, Eric. R., Schwartz, James H., Jessell, Thomas M. 2000).
Accumulation of sensory evidence in lateral intra-parietal (LIP) neurons#
As described before, the LIP is an oculomotor centre that is anatomically situated between the sensory (MT and MST) and motor areas (FEF, SC). Activity of a subset of neurons in LIP (Shadlen & Newsome 1996), are thought to integrate the sensory information of a stimulus, received via MT to reflect signatures of a decision process during a random dot motion (RDM) direction discrimination paradigm. (shown in figure 1.2 A). The task demands the monkey to make a choice between two different target locations based on the direction of randomly moving dots. Task difficulty depends upon the number of dots that move coherently in one direction. The key feature of the task is that it links decisions with the response of an eye movement. Thus, in this framework, problem of decision making could also be interpreted in the context of movement selection and preparation. MT neurons are not only tuned to the direction of moving dots but also have a persistent activity and its level depends upon the motion strength (inset for figure 1.2 B). When this information reaches LIP, a decision is said to be made when the population response reaches a fixed threshold. The rate at which the activity ramps up, is dependent on the motion strength of the stimulus and is thought to reflect the evolving decision process. As described before, these signals are then projected to higher oculomotor centres like the FEF and the SC to convert them into motor commands to initiate eye movements.
Additionally, micro-stimulation studies (Gold & Shadlen 2000, 2003) have revealed that the FEF is not only involved in response preparation but that the evolution of these decision variables happen very early in FEF - during the decision process itself. The leakage of decision signals from the LIP to the FEF could be tracked during an interrupted direction decision task (see figure 1.2 C). When the stimulation was done at different viewing durations, firstly the evoked saccades from FEF micro stimulation tended to deviate in the direction of choice made by the macaque (shown in figure 1.2 D) and secondly, the deviation was also dependent on the motion strength and viewing time (in figure 1.2 E). These results suggest that as the decision variables evolve over time in oculomotor structures like LIP, FEF and SC, information flows continuous from sensory neurons in MT to motor neurons in FEF and SC via LIP.
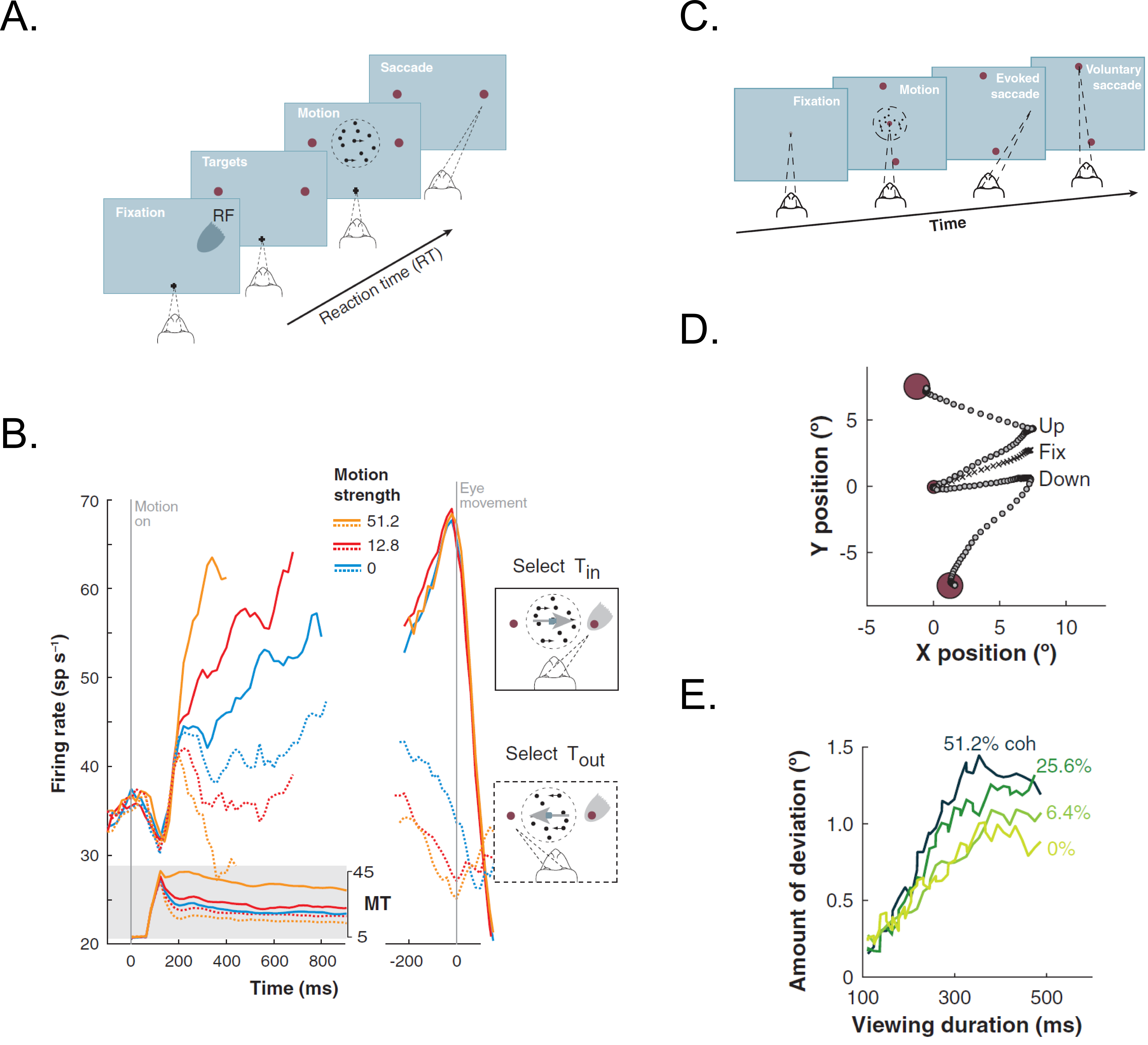
Figure 1.2 The neural basis for decision making and evolving decision variable in LIP neurons.#
A) Choice-reaction time version of the direction discrimination task. In this task, the macaques had to report the direction of the dynamically moving dots by making saccadic eye movements to the appropriate target location whenever ready. B) The average response of LIP neurons approximates the integral of a difference in firing rate between the MT neurons (shaded insert) with opposite direction preferences during the decision formation time. The indicated response was grouped based on motion strength and direction of choice. They are aligned on onset for random-dot motion (in left) and saccade onset (in right). C) Interrupted direction discrimination task. In this task, the motion viewing is interrupted by microstimulating FEF to evoke a saccade. Thus, the macaque had to make a second, voluntary saccadic eye movement to the target in periphery to indicate his decision. D) Example of eye movement trajectories when FEF was microstimulated during the fixation and while viewing upward and downward motion. The induced saccades deviated towards the direction of subsequent voluntary eye movements. E) The average amount of deviation depended on motion strength and viewing time. These results show that the information evolves over time in the oculomotor system and the outcome of the decision process. Figure taken from Gold & Shadlen 2007.
Motor planning in frontal eye fields (FEF)#
FEF forms the rostral bank of the arcuate sulcus in macaques. It is a critical node in the oculomotor system and is thought to play a major role in target selection and response preparation. Both these cognitive processes are essential for planning ‘where to’ and ‘when to’ make a goal directed saccade. Bruce & Goldberg 1985 recorded from a subset of neurons called visual and movement cells. Some of their properties are described below: Visual cells in FEF play an important role in target selection. Some of the neurons not only respond to an object in the receptive field but also are able to select a target among other distractors present in a visual scene. In an odd ball search task (Schall et al. 1995, Schall & Hanes 1998), multiple stimuli were presented in the visual field. Macaques were trained to make a saccadic eye movement to an odd ball target (red among greens or green among reds). The activity during the early transient phase in a visual cell was same for both the target and distractors in the receptive field. However, the activity during the later phase of a visual cell could be used to discriminate between the target and a distractor (shown as shaded area in figure 1.3 A). Such cells represent the goal or spatial location of an upcoming saccade in voluntary eye movements.
Movement neurons on the other hand, show increase in activity prior to a saccade and are known to play part in response preparation. The response of these cells does not depend on the presence or absence of a target at the saccade location and show tuning for different parameters like direction and amplitude of a saccade. Hanes & Schall 1996 showed that the time it took to initiate a saccade (also known as reaction time) could be associated to the rate at which the activity ramped up to reach a fixed threshold in movement cells, which was unique for each neuron and did not depend on reaction time. As shown in figure 1.3 B, the growth rate of the activity could predict the reaction time of an upcoming saccade. Steeper slopes are associated with shorter reaction times and vice versa. Similar neurons were also found in other regions of the brain like SC (Dorris et al. 1997) and LIP (Gold & Shadlen 2007, Roitman & Shadlen 2002). Furthermore, during a simple saccade task in some of the trials, a ‘STOP’ signal was randomly introduced after a certain delay (<120 ms) from time of target onset. The macaques had to successfully maintain fixation and cancel the saccade to get a reward. (Hanes et al. 1998) showed that the firing activity for movement neurons did not reach the threshold in those trials where the macaques were able to cancel an upcoming saccade (see figure 1.3 C). Together these studies suggested that the movement neurons do contribute to play an important role in initiating of a saccade. To summarise, FEF is thought to play a crucial role in sensory–motor transformations and have been shown to be involved in planning a goal directed saccade.
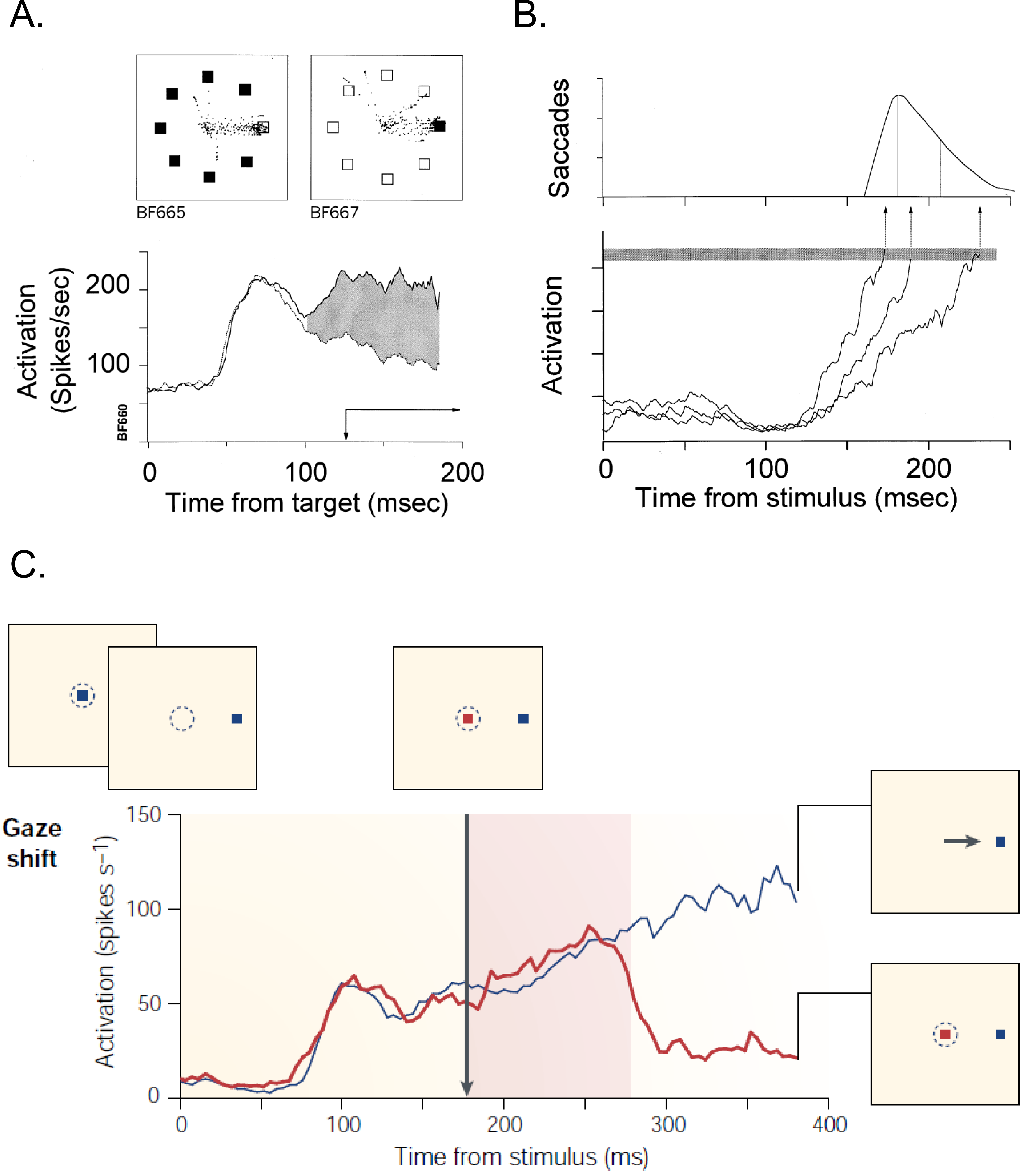
Figure 1.3 Neural basis for target selection, response preparation and control of purposeful eye movements.#
A) The time course of activation of a single FEF neuron, when the target (solid line) or a distractor (dotted line) was presented in the receptive field during a search task (as shown in panel above). The filled region indicates the interval when the two curves were significantly different. This result reveals that FEF plays an important role in target selection. B) The activity of movement-related neuron is related to the variability in saccade latency. It is accounted for by the time it takes to reach a fixed threshold.(Schall & Hanes 1998). C) Plots compare the rapid change in activation of neuron for different conditions by exerting control through cancelling a planned movement. (Schall 2001)
Linking cognitive processes with neural activity through accumulator models#
The characteristic pattern of activity seen in neurons of the LIP, FEF and SC, suggest that decision making and motor planning can be understood within a computational framework, provided by so called accumulator models, initially proposed by cognitive psychologists. These models characterise reaction time (RT) using different approaches and have been useful in understanding speed-accuracy trade-offs (Reddi & Carpenter 2000), response biases based on prior information (Carpenter & Williams 1995), the degree of urgency (Reddi & Carpenter 2000), competitive situations (Leach & Carpenter 2001), errors in decision making (Ratcliff 2001). In this framework, an internal signal rises to a certain criterion which marks the completion of a cognitive process or initiation of an action. The internal signal could reflect a decision variable (DV), state of the system, urgency or any other parameter relevant for the study. The simplest form of this model, which has been used in this thesis is the LATER (Linear Approach to Threshold with Ergodic Rate) model.
The LATER model here has been discussed in the context of a simple reaction time task, which is used for studying response preparation. The task demands to initiate a movement to a target or stimuli presented in the visual field. After a certain efferent delay, a preparatory signal starts from baseline (So) to rise linearly at a constant rate (r) towards a fixed threshold (ST). Once the criterion or the threshold is reached, the system is prepared to make a response. This model captures the variability in reaction time distribution across trials using only 4 parameters (i) baseline, (ii) slope or rate of rise, (iii) fixed criterion or threshold and (iv) onsets (as shown in figure 1.4 A). In the model, constant ‘r’ varies from trial to trial with a gaussian distribution which has a mean mu (μ) and variance sigma (σ). The distance (θ) between the baseline and threshold remains fixed across trials.
For a fixed threshold model, changes in the different parameters lead to systematic changes in reaction time (as shown in figure 1.4 B). With increase in baseline activity (So), the average reaction time and the variability for the distribution decreases. Similarly, for steeper slopes (r), the mean as well as the variability of the reaction time distribution decreases. Also, with delays in onset conditions, average reaction time increases but however, the variability for the distribution remains unaffected. Thus, systemic changes in RT can be attributed to changes in one or more of these parameters that describe the LATER model. In addition, LATER models have been extended to incorporate race models with two or multiple competing accumulators, which can explain behaviour in countermanding and redirect tasks (Camalier et al. 2007, Hanes & Carpenter 1999, Kapoor & Murthy 2008), gap paradigms (Story & Carpenter 2009), GO/NO-GO tasks (Noorani et al. 2011) and anti-saccade tasks (Noorani & Carpenter 2013).
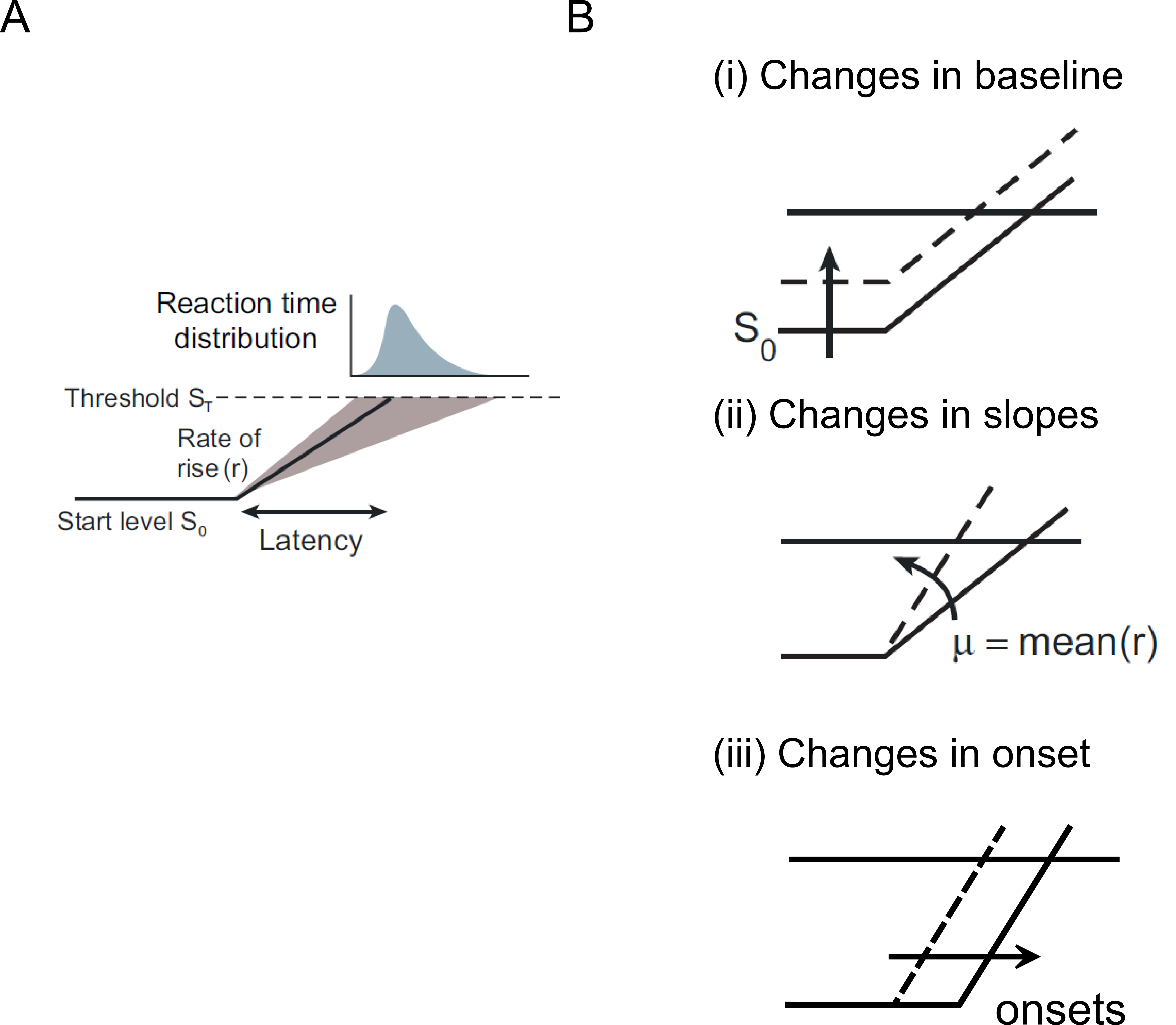
Figure 1.4 The LATER model.#
A) Response of the system is initiated when a DV or the preparatory signal reaches a fixed threshold (ST). The rate at which the signal rises (r) is predictive of reaction times. The distribution of predicted RT is skewed. B) The model predicts systematic changes in reaction time with changes in (i) baseline, (ii) slope and (iii) onsets. (Figure taken and modified from Gold & Shadlen 2007).
Movement preparation in dorsal premotor cortex (PMd)#
The accumulator model is a one-dimensional (1D) model and has been used extensively in the oculomotor system to study decision-making and motor planning of saccadic eye movements. However for the hand movement planning, Shenoy and colleagues suggested a more dynamic view to study movement preparation in dorsal premotor area (Shenoy et al. 2011). Based on their experiments that involved recording from multiple neurons simultaneously, they proposed that the variability in the neuronal response of dorsal premotor cortex (PMd), in a multi-dimensional subspace could predict the RT for initiating hand movements. They called this as a subspace hypothesis (Afshar et al. 2011, Churchland et al. 2006, 2010; Kaufman et al. 2014). In a N-dimensional neural space, during the preparatory phase the motor system tries to optimise the configuration of the firing pattern of neurons to lie within a manifold or a subspace for initiating desired movements. Longer trajectories and the preparatory state that lies closer to the movement space are associated with longer reaction times. Similar to the diffusion process the stochastic trajectory of the state space also predicts the reaction time but in a multi-dimensional space. Indeed, a highly reduced version of their state space model, equivalent to the one-dimensional accumulator model, did a reasonable job of predicting hand movement RTs, as well. In this context, it is interesting to note that ramping up activity during movement preparation is also observed in scalp electrodes that reflect global correlates of motor planning Gratton et al. 1988. Moreover, ramping up activity during movement preparation have also been reported for wrist movements (Riehle & Requin 1993), suggesting the applicability of accumulation to a fixed bound as a reasonable first approximation to study motor planning and preparation for hand movements as well.
Separating motor plans from execution#
As alluded to in the initial paragraph, in a fast-changing environment, situations and circumstances might demand us to have a separate planning and execution stages. A similar separation of motor planning and execution is also implemented by our saccadic system, to allow flexible control of eye movements to ensure that saccadic plans are countermanded if necessary (Hanes et al. 1998, Schall 2001). Sometimes it is best to change our decisions based on our confidence levels as well (Kiani & Shadlen 2009).
In all the examples illustrated above, even though the information required to make movements was being processed by our central nervous system it did not lead to any overt action. In fact, our brain might be spending most of its energy in developing different computational architectures and/or allocating resources to separate these different mental processes from being executed and merge them based on task context, if necessary. This gating of information between intention and action could happen at different levels in our central nervous system involving cortical and sub-cortical structures like the motor cortex, basal ganglia and even the spinal cord.
Cortical and Sub cortical structures that gate information between planning and execution stages#
The traditional view is that different cortical structures like FEF (part of oculomotor network), PMd and M1 (part of skeletal motor system) play an important role in planning and initiating eye and hand movements, respectively. Recent studies suggest that both these areas also have gating mechanisms (Eye: Boucher et al. 2007, Hanes et al. 1998, Schall et al. 2000, Schall 2001, 2004 ; Hand: Gallego et al. 2017, Kaufman et al. 2014) to separate motor plans from being executed. These gates could be computational or structural.
Computational gating mechanisms in FEF and PMd#
A subclass of neurons called the fixation neurons have also been identified and studied in FEF. They are tonically active during the fixation period. Prior to and during a saccade, the activity gets suppressed and stays low throughout. However, just before the saccade terminates the tonic activity of these fixation neurons get restored. Flexibility in behaviour requires that one can stop a saccadic plan depending upon different situations. Towards this goal, studies have shown that after a stop signal appears during the planning stage the activity in fixation neurons increase rapidly. In contrast, the activity of a movement neuron decreases after the stop command appears. Neurons associated with inhibitory processes are not just present in the FEF, but have also been identified within the rostral pole of the SC, where they are called fixation neurons and are active during fixation, but decrease their activity prior to saccades. Such fixation neurons are thought to be connected to a group of neurons in the brainstem called omnipause neurons which inhibit the burst neurons that are responsible for the rapid muscle contraction that occurs during saccades. Thus, in additions to neurons planning saccades there is a dedicated circuit of interconnected neurons that are responsible for preventing or inhibiting motor plans from prematurely getting executed.
Shenoy and colleagues more recently suggested that the motor cortex could act like a filter, allowing only certain pattern of activity in the cortical neurons to affect appropriate muscle groups. During the preparatory phase, the cortical activity lies within a manifold in a N-dimensional neural space (each dimension represents activity of a single neuron). They observed that the output of the population response that could lead to muscle contraction lied in the potent dimension or outside this manifold. They also observed that even though the neurons were individually involved in movement preparation but if the firing pattern of the population fell inside the manifold, the activity could cancel out each other and hence show no effect on muscle groups that lie downstream to PMd and is connected via spinal motor neurons (see figure 1.5 a,b). They call this manifold as the null space.
Basal ganglia as a structural gate#
Sub-cortical structures like the basal ganglia also play an important role in movement initiation. The BG is conceptually thought to act like a structural gate, keeping all the potential signals from cortex at check until it provides a triggering response to SC or thalamus to initiate eye and hand movements, respectively. If the gate is open, it allows the information from different cortical regions to reach appropriate muscle groups via its downstream effectors. However, if the gate is closed, the pattern of activity in cortex does not matter, the information cannot reach to its targeted downstream areas (see figure 1.5 c,d). The basal ganglia select appropriate and inhibits inappropriate movement plans by exerting strong tonic inhibition via its two output nuclie: substantia nigra pars reticulate (SNr) and globus pallidus interna (GPi). It receives information from different cortical areas like FEF and somatosensory areas that reach caudate and putamen. Together these input structures are called the striatum. The caudate nuclie projects inhibitory signals to SNr. Thus, when movement plans from cortical areas reach caudate, they inhibit SNr. The inhibition on SNr disinhibits the tonic inhibition on SC. Thus, allowing activity in SC to build up for initiating eye movements.
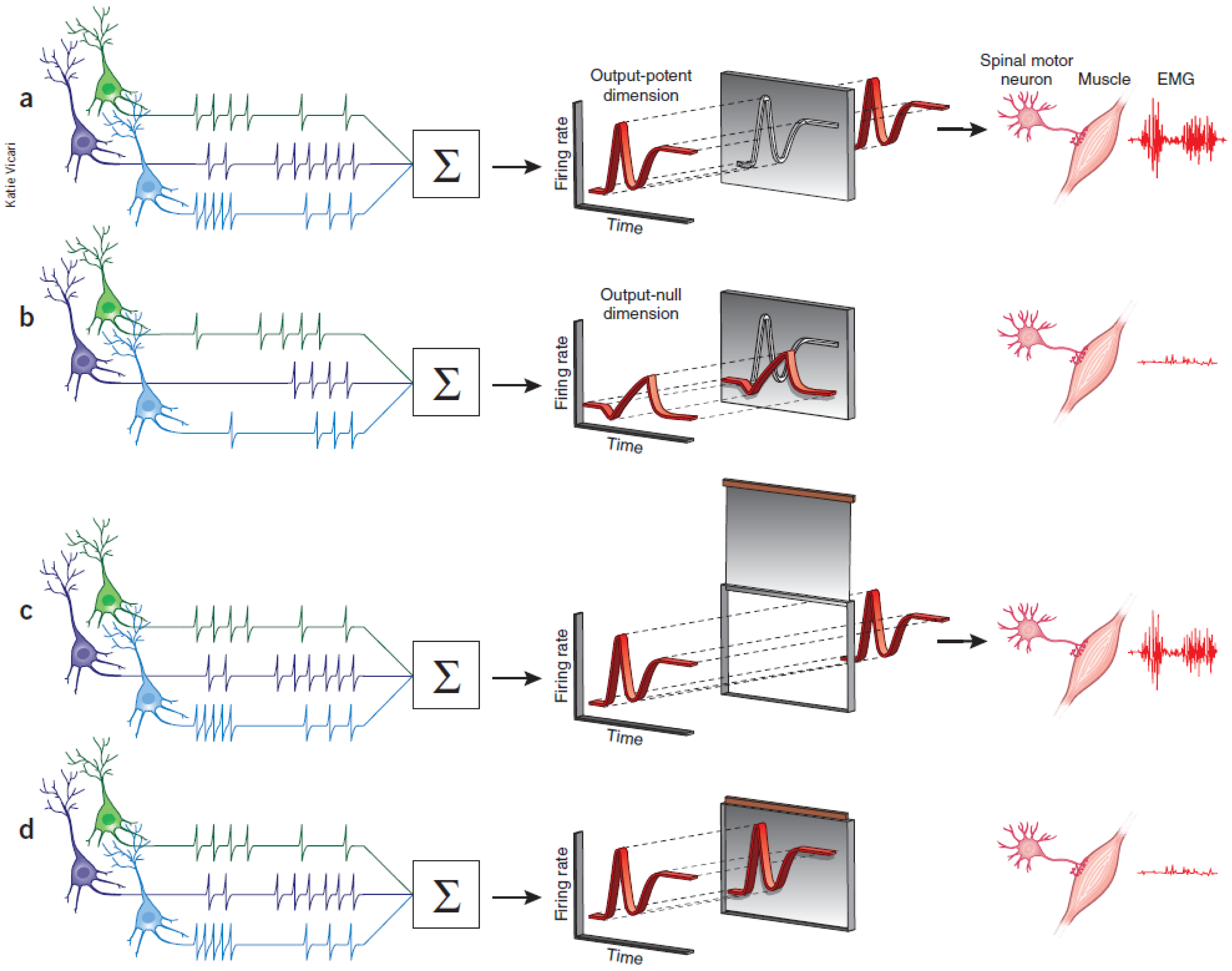
Figure 1.5 Gating mechanisms for flow of information into central nervous system.#
A highly schematic illustration of computational (a,b) and structural (c,d) architectures that could gate the flow of information in different cortical and sub-cortical areas. (a) Only certain patterns of motor cortical activity effect appropriate muscle groups (b) Different patterns of descending cortical activity cancel out and have no effect on downstream effectors. The cortical areas can act as a filter allowing only some pattern of neural activity that lie in an output-potent dimension to reach the periphery. (c) In the motor gating hypothesis, irrespective of the firing pattern of different input areas, if the structural gate is open, motor commands reach appropriate muscle groups. (d) Similarly, the neural activity cannot affect the muscle group if the structural gate is closed. Figure was taken from Sanger & Kalaska 2014.
Spinal Cord#
Despite the gating afforded by cortical and sub cortical structures, Prut & Fetz 1999 showed that preparatory changes during the delay period could be tracked down till the level of spinal interneurons. The activity during the delay period reflected movement planning as they were correlated to different parameters like direction and extent of an upcoming movement. Their work also suggested that both excitatory and inhibitory processing occur during the delay period at the level of spinal circuitry. They recorded activity from spinal interneurons while macaques were trained on a flexion/extension task with an instructed delay period (See figure 1.6 A). The amount of force that was applied on a fixed manipulandum was mapped on to cursor movements on the screen. In short, the monkey had to maintain the cursor within the central spot to start a trial. The delay period separated ‘where’ from ‘when’ to move the cursor to the filled target location. After the go cue, the monkey had to move and hold the cursor at the target location to get a juice reward. Most of the spinal interneurons showed a decrease in activity from baseline during the delay period.
Furthermore, recent studies (Cohen et al. 2010, Prut & Fetz 1999, Sinclair & Hammond 2009) suggest that both excitatory and inhibitory processes occur during the delay period at the level of spinal circuitry. These two competing processes are also known as the ‘priming and braking’ mechanism. The cortico-spinal (CS) tracts exert an excitatory influence on spinal interneurons and they carry task relevant information from premotor areas (PM) to motor cortex (M1) to spinal cord, thereby priming the interneurons for movements during the preparatory phase. Also, many of the spinal interneurons receive strong inhibitory signals from premotor cortex via the reticulate formation. Cohen et al. 2010, suggested that since the premotor areas are dominantly active during the preparatory phase; these interneurons may reflect the superimposed global inhibition which suppresses the tendency to initiate movements due to priming. Prior to a movement the activity of premotor area decreases, which lifts the global inhibition in the circuitry. Thus, revealing the already primed circuitry to effect appropriate muscle groups.
Such mechanisms in spinal cord might act like a gate, not allowing preparatory signals to reach appropriate muscle groups during the delay period. Prut & Fetz 1999 also recorded from distal muscle groups while the macaques were performing an instructed delay time task. No significant changes were observed in the muscle activity during the delay period. Example of muscle recordings from a trial during one of the sessions is shown in figure 1.6 A. Modulations were seen only after the go cue, in extensor digitorium muscles (blue trace) and not in Flexor digitorium superficialis (red trace). As the name suggests only, the muscle which were required to apply the force in appropriate direction was active. Their findings suggest that even though preparatory changes could be detected at the level of spinal cord, information about the upcoming movement was gated out at the level of spinal cord and did not leak through into the periphery during the delay period.
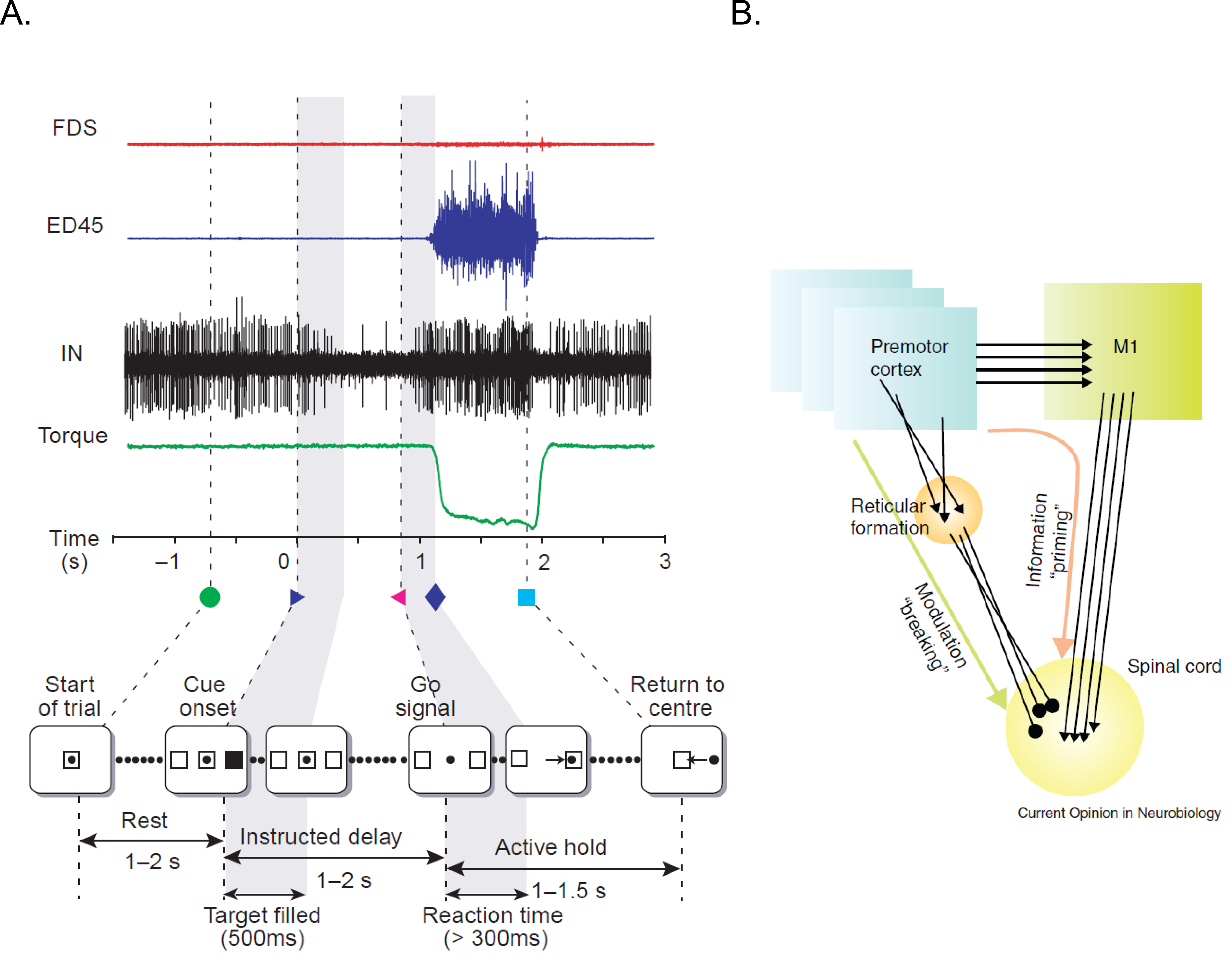
Figure 1.6 Processing of information at the level of spinal circuitry.#
A) Example of a single instructed delay trial is shown at the bottom panel. Traces (from top) show activity of two different muscle groups (Top: flexor digitorum sublimis and extensor digitorum). Shown also is the activity of interneurons and torque signal. During the delay period, the firing rate of this unit decreases, with no accompanying torque deflection or EMG activity of either muscle. B) The illustration represents a possible model to account for the priming and braking mechanisms in spinal cord circuitry during movement preparation. Figures have been taken from Prut & Fetz 1999 and Cohen et al. 2010.
Leakage of information into the periphery#
Despite the presence of brain structures, circuits and computational mechanisms that gate information flow between central and peripheral structures behavioural paradigms such as visually guided movements and inhibition of return in cueing paradigms have shown that recruitment pattern for neck muscles parallel the activity of neurons in intermediate and deep layers of SC (Corneil et al. 2008). Task related information (example, pro or anti saccade) and stimulus driven responses do manifest themselves in peripheral muscle groups. Anti-saccade tasks have revealed that both the top down and bottom up influences affect the recruitment of neck muscles in the periphery (Chapman & Corneil 2011). Signatures of other cognitive process such as motor preparation (Corneil et al. 2007, Dorris & Munoz 1998) and impact due to reward (Roesch & Olson 2003) have been recorded from neck muscles as well.
In the context of hand movements, many studies such as Georgopoulos et al. 1988, Lecas et al. 1986, Tanji et al. 1988, consistent with Prut & Fetz 1999, suggest that the information about the upcoming movement gets gated out at the level of spinal cord and does not leak through into the periphery. However, there are a few studies that suggest otherwise. Studies done by Mellah et al. 1990 and Duclos et al. 2008, show that motor units do get recruited during the delay period and there are changes in the tonic firing pattern during the preparatory phase. Furthermore, both these studies reflect excitatory and inhibitory processes in spinal cord.